多日重新学习一项自动技能。(a)自动抓取设置。门打开后(通过声音发出信号),老鼠从一个狭缝中取出小球。插图中的M1(绿色)和DLS(灰色)位置。(b)再学习范式。老鼠被过度训练以到达位置A(左)。然后小球移动到位置B继续训练(中,右)。(c)第一终点。上图:例子动物,端点x位置直方图,横跨试验。底部:相对于小球位置(大黑圈)的示例到达轨迹和端点位置(与顶部相同)。 (d) Decay in reaches to A. Percentage of trials in a session with first reach (R1) to location A, as compared to low-amplitude (short, S) reaches and/or reaches to location B. (e) For those trials with multiple reaches, within-trial updating for reaches after the first were classified into short (S), old (A), or new (B) reaches. (f) Example x-trajectory of paw during a trial, with reach onset marked; r1 and r2 are the first and second reach onsets, respectively. g Reach type after first reach, for trials with multiple reaches. For all reaches after the first reach in a trial, proportion of A, B, or S reaches. Data are presented as mean values ± SEM. h Following an inaccurate trial, we classified the first reach type of the subsequent trial. i Reach type after inaccurate reach to A on previous trial. Data are presented as mean values ± SEM. *<0.05; **<0.01; ***<0.001, ANOVA, Tukey-Kramer post hoc test. Credit:自然通讯(2022)。DOI: 10.1038 / s41467 - 022 - 30069 - 1
神经科学家的目标是了解初级运动皮层(M1)可以调节精确的运动,同时支持遇到一致错误的行为探索。虽然环境的一致性可以帮助动物学习和自动化高度可靠的行动,但它们必须保持灵活性以适应环境的变化。在一份新报告中自然通讯美国加州大学旧金山分校(University of California, San Francisco)的Sravani Kondapavulur和一个神经工程研究团队,在大鼠身上开发了一种伸手抓握的任务,以监测初级运动皮层(M1)和神经的可变性背外侧纹状体(DLS)地区。结果表明,与行为探索中观察到的可变神经模式相比,运动区和纹状体区如何通过生成自动和精确运动的可靠神经模式在两种模式之间过渡。
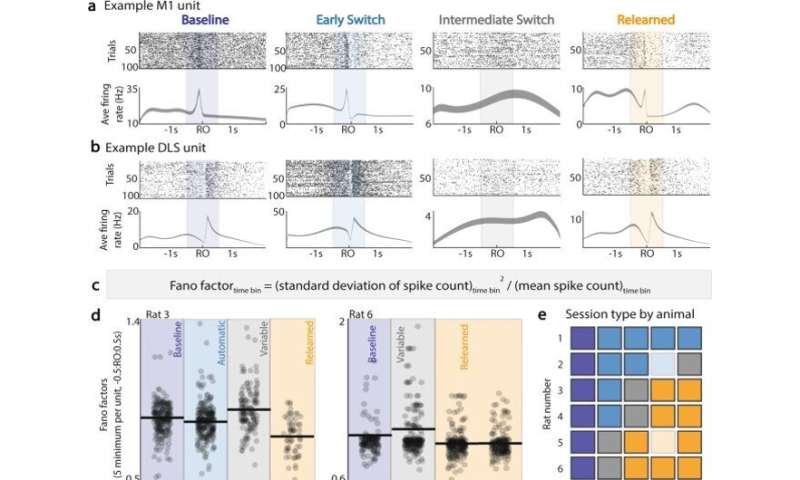
在勘探过程中,M1和DLS单元被锁定。(a)顶部:单个M1单元†跨时段的栅格图(跨天的同一频道)。下:试验平均射速(平均值±SEM)在第一次到达后- 2秒至2秒内。(b)与(a)项相同,适用于临时登记单位。†(c)为每个单元,每个会话的每个时间仓确定范诺因子的方程。(d)对于两个例子动物,在第一次到达发病前后的−0.5 s至0.5 s内,每单位5个最小范诺因子。(e)基于Fano因子的会话类型重新分类:靛蓝色=基线,蓝色=自动(切换后,没有Fano因子变化),灰色=变量(Fano因子最大增加),橙色=重新学习。浅蓝色/浅橙色=由于记录过程中数据损坏,没有分析两个会话。(f)首次达到A的试验(蓝点)和首次达到B的试验(橙点)的M1单位峰值试验平均调制,跨会话类型;在z-计分单元在整个会话中发射后计算峰值调制。 (g) Same as (f), for DLS units. (h) Change in proportion of first reach to A in a session, relative to proportion of first reach to A in previous session; grouped by session type across all animals, all sessions. i Proportion of first reach to B in a session, grouped by session type across all animals, all sessions. †Unit may not be the same across sessions. *<0.05, **<0.01, ***<0.001, linear mixed effects model, Bonferroni-corrected significance. Credit:自然通讯(2022)。DOI: 10.1038 / s41467 - 022 - 30069 - 1
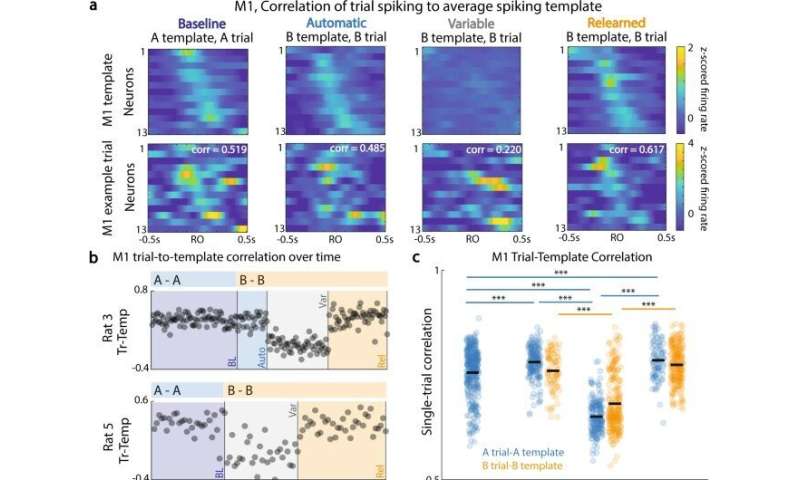
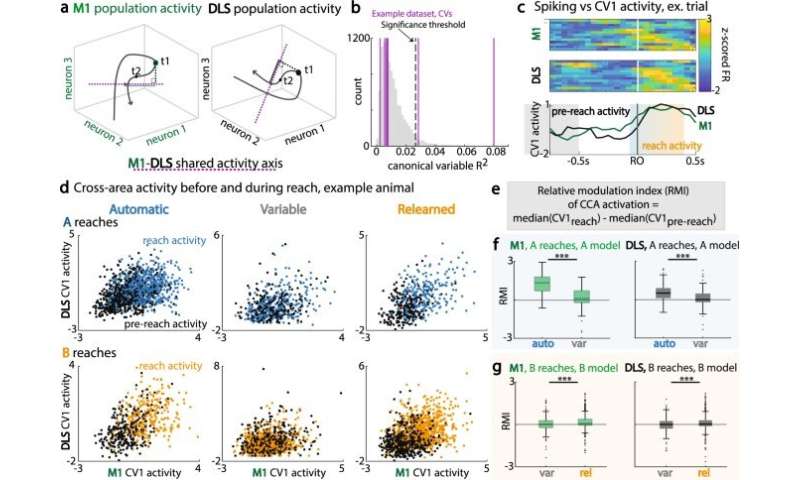
M1-DLS跨区域子空间调制的变化。(a)典型相关分析(CCA):确定M1和DLS活动(紫色虚线)之间具有最大相关性的轴,以及M1群体活动(左,绿色)和DLS群体活动(右,黑色)在t1时刻的预测群体活动的示例值(共享轴上的实线)。(b)用于识别重要规范变量(CV)轴的R2值的自举洗牌分布示例。(c)上:单次试验M1峰值活性。中:单次试验DLS峰值活动。底部:沿着CV1对应的单次试验M1(绿色)和DLS(黑色)激活。预到达周期(灰色)是第一次到达开始前的−1到−0.5秒。到达周期(蓝橙色)为−0.1 s至0.4 s左右首次到达。(d)到达前(黑色)和到达期间(彩色)M1和DLS跨区域活动,对于跨会话类型的A试验(上行)和B试验(下行),例如动物。(e) CCA激活的相对调制指数(RMI)方程,比较到达周期和到达前周期。 (f) RMI for trials with first reach to A, for M1 activations (left, green) and DLS activations (right, green), across automatic and variable sessions. R1 to A, M1 and DLS RMI: auto, n = 220 trials; var, n = 95 trials. Data are presented as box plots with 25th, 50th, and 75th percentiles. (g) Same as (f), for trials with first reach to B, across variable and relearned sessions. R1 to B, M1 and DLS RMI: var, n = 150 trials; rel, n = 464 trials. Data are presented as box plots with 25th, 50th, and 75th percentiles. *<0.05; **<0.01; ***<0.001, linear mixed effects model, Bonferroni-corrected significance. Credit:自然通讯(2022)。DOI: 10.1038 / s41467 - 022 - 30069 - 1
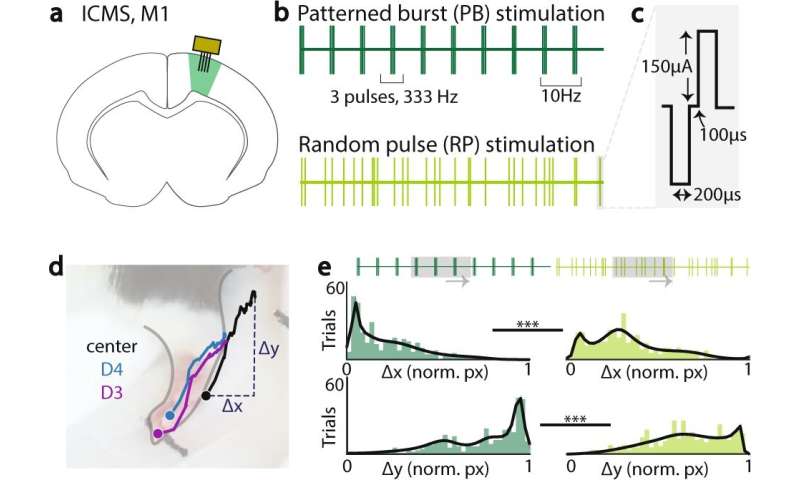
刺激变异性在驱动上肢运动变异性中的作用。a刺激的位置,通过微线阵列到M1,以识别引起前肢运动的区域。b增产方式,总电荷平衡。模式脉冲刺激(顶部,PB)由333 Hz三组组成,频率为10 Hz三组。随机脉冲刺激(底部,RP)由单脉冲组成,随机定时为30hz。每次刺激1秒,每次共30次脉冲。c刺激脉冲参数,采用200 μA双相脉冲,每相200 μs,相间间隔100 μs。d脚掌侧位图:标记用于监督运动跟踪的肢体标记:数字4的尖端(D4,蓝色),数字3的尖端(D3,紫色),脚掌中心(中心,黑色)。每个试验的终点位置(Δx, Δy)相对于试验窗口中的起始位置计算,并在不同刺激条件下由动物归一化。e范例动物直方图的归一化Δx(上)和Δy(下)的爪子中心在模式突发刺激试验(左)和随机脉冲刺激试验(右)。 Gray boxes show an example 300 ms window with identical number of pulses. Kolmogorov-Smirnov test for two samples in this example animal, center of paw, p = 1.8e−11; *<0.05; **<0.01; ***<0.001, Kolmogorov-Smirnov test for two samples. Credit:自然通讯(2022)。DOI: 10.1038 / s41467 - 022 - 30069 - 1
 Automated reach-to-grasp setup. After the door opens (signaled by a tone), the rat reaches through a slit to retrieve the pellet. M1 (green) and DLS (gray) locations in inset. (b) Relearning paradigm. Rats are overtrained to reach to location A (left). Then pellet moved to location B with continued training (middle, right). (c) Endpoint of first reaches. Top: example animal, histogram of endpoint x-position, across trials. Bottom: example reach trajectories and endpoint locations relative to pellet location (large black circle) for same sessions as (top). (d) Decay in reaches to A. Percentage of trials in a session with first reach (R1) to location A, as compared to low-amplitude (short, S) reaches and/or reaches to location B. (e) For those trials with multiple reaches, within-trial updating for reaches after the first were classified into short (S), old (A), or new (B) reaches. (f) Example x-trajectory of paw during a trial, with reach onset marked; r1 and r2 are the first and second reach onsets, respectively. g Reach type after first reach, for trials with multiple reaches. For all reaches after the first reach in a trial, proportion of A, B, or S reaches. Data are presented as mean values ± SEM. h Following an inaccurate trial, we classified the first reach type of the subsequent trial. i Reach type after inaccurate reach to A on previous trial. Data are presented as mean values ± SEM. *<0.05; **<0.01; ***<0.001, ANOVA, Tukey-Kramer post hoc test. Credit: <i>Nature Communications</i> (2022). DOI: 10.1038/s41467-022-30069-1")