CA1神经元在时间等分任务中的可扩展时间表示。(A)任务示意图(任务1)。让大鼠在跑步机上长时间或短时间跑步,然后分别选择长时间或短时间与水奖励相关的左臂或右臂。第1组和第3组的间隔是10秒(长)和5秒(短),第2组的间隔是20秒(长)和10秒(短)。(B, a到c)块1(左)、块2(中)、块3(右)三个代表性时间单元的放电模式。上图:长间隔和短间隔试验的栅格图。底部:长间隔(实线)和短间隔(虚线)试验的PETHs。(C)第1块(左)、第2块(中)和第3块(右)的时间细胞放电模式(n =来自7只大鼠的454个单元)。每一行表示每个块中单个神经元在长间隔试验中的peth。颜色刻度代表每个神经元的放电速率。神经元按照第1块中的峰值时间排序。 (D) Population vector analyses of the data (C). Left: Autocorrelation of the population vector matrix of block 1. Center and right: Cross-correlations of blocks 1 and 2 and blocks 1 and 3, respectively. The color scale represents Pearson’s correlation coefficient. (E) Distributions of the scaling factors of time cells between blocks 1 and 2 (blue) and blocks 1 and 3 (green). Credit: Science Advances, doi:10.1126/sciadv.abd7013
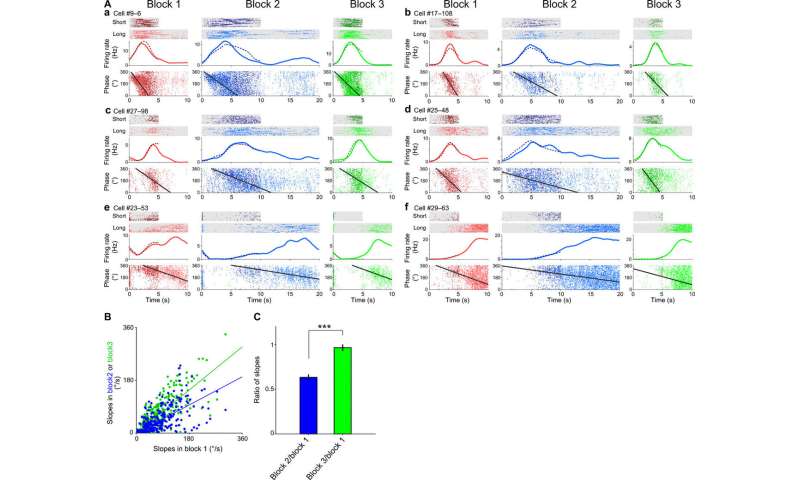
时间单元相位进动的可扩展性。(A, A to f)任务1中block 1(左列),block 2(中列),block 3(右列)中三个代表性时间单元的发射速率和θ相。上图:短间隔和长间隔试验的栅格图。中间:长间隔(实线)和短间隔(虚线)试验的PETHs。底部:相位图作为运行时间的函数。每个点代表长(浅色)和短(深色)间隔试验的动作电位。每个面板中的黑线表示数据的圆形-线性拟合(见材料和方法)。(B)任务1中每个单元(n = 351个单元)中块1和块2(蓝点)之间以及块1和块3(绿点)之间相位进动斜率的关系。实线表示这些数据的线性回归拟合。(C) block 2中每个cell的相位进动斜率除以block 1中的相位进动斜率(蓝色)。 Slope of phase precession in each cell in block 3 divided by that in block 1 (green). Means ± SEM. ***P < 0.001, paired t test. Credit: Science Advances, doi:10.1126/sciadv.abd7013
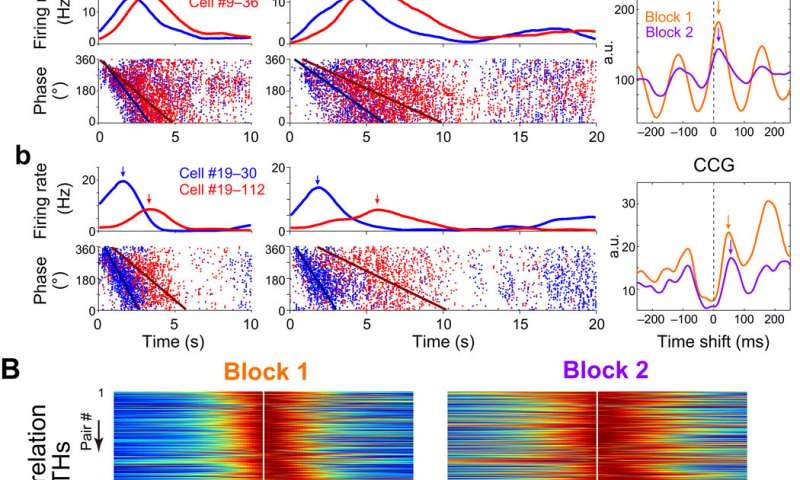
单θ周期内尖峰序列的时间压缩的两两分析。(A, A, b)左:第1块和第2块(任务1)中两对具有代表性的时间单元(红色和蓝色)的发射速率和θ相。箭头表示发射速率峰值。右图:第1块(橙色)和第2块(紫色)中对的交叉相关图(CCGs)。箭头表示ccg的峰值。请注意,(Ab)的CCG峰值时移比(Aa)(右)的更大,反映了(Ab)的PETH峰值时间比(Aa)(左)的更大距离。A.u,任意单位。(B)时间单元对peth的相互相关性。(C)第1块和第2块成对时间细胞的ccg (n = 694对)。在(B)和(C)中,对的顺序相同。(D)在区块1和区块2中,对单theta周期中spike序列的时间压缩进行成对分析。每个点代表一个神经元对。 Horizontal axes show peak distances of PETHs, and vertical axes show peak shifts of the CCGs. The slopes of the regression line (red) were 29.2 and 15.0 ms/s in blocks 1 and 2, respectively. (E) Relationship between the time shifts of the CCGs from blocks 1 and 2. The slope of the regression line (red) was 0.98. Credit: Science Advances, doi:10.1126/sciadv.abd7013
 Schematic of the task (task 1). The rats ran on the treadmill for long or short intervals and then selected the left or right arm associated with water reward for long or short intervals, respectively. Sets of intervals were 10 s (long) and 5 s (short) in blocks 1 and 3 and 20 s (long) and 10 s (short) in block 2. (B, a to c) Firing patterns of three representative time cells in blocks 1 (left), 2 (center), and 3 (right). Top: Raster plots of long- and short-interval trials. Bottom: PETHs of long (solid)– and short (dotted line)–interval trials. (C) Firing patterns of time cells in blocks 1 (left), 2 (center), and 3 (right) (n = 454 units from seven rats). Each row represents PETHs of single neurons during long-interval trials in each block. The color scale represents the firing rate of each neuron. The neurons were ordered according to the peak time in block 1. (D) Population vector analyses of the data (C). Left: Autocorrelation of the population vector matrix of block 1. Center and right: Cross-correlations of blocks 1 and 2 and blocks 1 and 3, respectively. The color scale represents Pearson’s correlation coefficient. (E) Distributions of the scaling factors of time cells between blocks 1 and 2 (blue) and blocks 1 and 3 (green). Credit: Science Advances, doi:10.1126/sciadv.abd7013")








用户评论