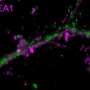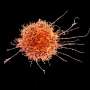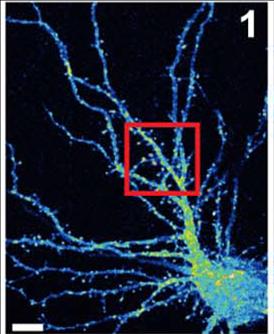
一个老鼠海马神经元(细胞体在右下)的伪彩色图像,它的树枝状树突提供了表面,其他神经元的投射物可以通过形成突触连接起来。Van Aelst和同事们已经证明,当OPHN1蛋白发生突变,干扰了它与另一种名为Homer1b/c的蛋白质相互作用的能力时,AMPA受体不会以通常的速度在突触上循环到表面。这对突触可塑性产生了负面影响,而突触可塑性是神经元调节连接强度的过程。这种病理可能在x -连锁智力迟钝中起作用。资料来源:CSHL范·埃尔斯特实验室
有新的线索表明,大脑细胞的功能异常会导致智力残疾,可能还会导致其他发育性大脑疾病。
冷泉港实验室(CSHL)的Linda Van Aelst教授一直在仔细研究一种名为OPHN1的正常蛋白是如何帮助大脑兴奋性神经传递的,特别是在含有AMPA受体(AMPARs)的神经细胞对接口。她的团队的新工作,发表于6月24日神经科学杂志》上该研究为OPHN1缺陷如何导致ampar表达神经元突触强度成熟和调整的障碍提供了新的机制。ampar表达神经元普遍存在于大脑中,并对兴奋性神经递质谷氨酸作出反应。
位于X染色体上的一种名为寡分裂蛋白1 (OPHN1)的基因的突变,此前曾被认为与X染色体相连智力障碍(也被称为X-linked mental retardation)是一种严重影响男孩的疾病,在所有的男性智力残疾中,这种疾病可能占到五分之一。
到目前为止,人们已经发现了几个不同的OPHN1基因突变,所有这些突变都会扰乱神经细胞对OPHN1蛋白的制造。此前,Van Aelst和他的同事证明了OPHN1在突触可塑性(邻近神经细胞调节其连接强度的过程)中起着至关重要的作用。当大脑细胞对刺激流做出反应时,它们会不断调整连接强度。
这项新发现表明了OPHN1是如何参与ampar的传输的,这是神经元可塑性的一个基本特征。神经元将受体从突触转移到突触内部,然后再返回到突触表面以控制连接强度。在突触表面,受体为神经递质的对接提供了机会,在这里是谷氨酸分子。当一个细胞被激活后,表面的受体通常会被带回到内部,在那里它们被循环利用以备将来使用。
当OPHN1由于基因突变而畸形或缺失时,CSHL团队证明,它不能再正常地发挥受体循环的作用,从而也损害了神经元与邻近神经元保持长期紧密联系的能力,这被称为长期增强。
Van Aelst的新实验解释了与另一种名为Homer1b/c的蛋白质复合物中的OPHN1通常如何与内吞区(EZ)相互作用,以提供一池ampar,并将其带到突触后密度(PSD)位置。当OPHN1发生突变时,神经池不会形成,强化突触所需的受体也没有。长期增强功能受损。
范·埃尔斯特博士说:“这表明了一种以前未知的方式,即OPHN1基因缺陷会导致谷氨酸系统功能障碍。”“我们早期的研究已经表明,OPHN1对于稳定突触上的AMPA受体至关重要。这两种重要作用共同表明,有缺陷的OPHN1蛋白可能会导致导致x -连锁智力残疾的病理。”
期刊信息:神经科学杂志》上
所提供的冷泉港实验室