2020年2月12日报告
用多巴胺氧化还原穿梭器编程电子生物计算机

尽管大多数神经递质发挥着相同的基本作用,但它们是不可替代的。每一种都从传入的动作电位中接过接力棒,并将神经信息通过突触分裂传递,但每种类型的传递器都有自己独特的扭曲。简单地说,它们有完全不同的代谢作用。正常运转的大脑需要所有这些器官的兼收并蓄的服务,无论何时何地。几乎在大脑的每一个偏远区域,无论是视网膜、嗅球,甚至是脑干的特定发射器投射核,都可以在其中找到所有主要的发射器系统。
帕金森氏症是多巴胺能传递的毁灭性失败。只要看看病变的基底神经节——一个很大程度上建立在多巴胺基础上的整个神经生态系统——就能看到对其回路的任何重大扰动所造成的毁灭性影响。然而,尽管有无数的研究,我们仍然不知道多巴胺对大脑到底有什么作用,更不用说对单个突触了。换句话说,是什么让它不可替代?
如果它们被适当地利用,危险的反应在生物学中也可以是有用的反应。关于为什么多巴胺神经元特别容易受到氧化损伤,已经有很多议论。通常认为线粒体和氧化还原反应是罪魁祸首。然而,一连串的事件紧密相连突触传递对那些有力量的过程来说,它还没有出现。
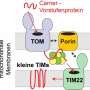
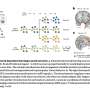
在这一空白中,最近发表在自然神经科学电子传递链.为了做到这一点,电子必须以某种方式穿过外膜,然后穿过膜间空间,最终进入内膜室。它在共价结合的黄素辅因子FAD的帮助下完成这一过程,FAD在此过程中转化为FADH。FAD可以以四种氧化还原状态存在,是电子和质子的多功能穿梭器。
为了再生,FADH被认为在细胞质中与分子O2反应形成FAD和过氧化氢。然而,作者没有发现H2O2利用他们的细胞质传感器。在一个附带的评论,建议H2O2可以以某种方式定位于线粒体,并可以将电子捐赠给膜间空间中的可溶性受体。从那里,他们可以转移到分子氧在呼吸复合体IV。
原论文的通讯作者Jim Surmeier向我提出这个膜间空间载体可能是细胞色素c,但没有提到H2O2具体来说。2O2?从这些结果中显然很难看出这一点。
为了更好地处理桌子上的许多种电子载体,在这里,我们将快速看一看略有不同的内膜酶系统,琥珀酸脱氢酶或呼吸复合物II。琥珀酸脱氢酶也将FAD转化为FADH,但它立即将电子通过铁硫簇转移到泛醌(Q)上形成QH2.这么多2然后将电子摆渡到复合体III。这是我们更熟悉的方法。复合体I做了类似的事情,只是它在生成QH之前的初始步骤中使用了NAD2.事实上,其他几个关键的内膜蛋白,包括DHODH(二氢磷酸脱氢酶),ETF(电子转移黄蛋白)和GPDH(甘油-3-磷酸脱氢酶),都使用可溶性QH将电子发送到复合体III2母舰。
在他们的实验中,作者通过阻断复合物III使所有这些电子通路短路。他们还阻断了腺嘌呤核苷酸转运体,以防止细胞质ATP反向驱动复合体V。通过这些操作,他们发现多巴胺能够恢复线粒体内膜上的电势。只有在提供多种刺激以引起持续的活动后,ATP水平才会下降多巴胺的释放被削弱。
虽然有可能,但还有许多其他东西可以控制多巴胺的水平,就此而言,任何传递器都可以。更一般地说,所有突触都倾向于遵循一个非常基本的操作计划的某种变体。首先,你向突触间隙释放大量的递质。然后会发生两种情况中的一种:要么递质被突触外的酶修饰——突触前位点和突触后位点都在争夺这些酶的好处——要么这两个位点立即重新摄取未修饰的递质,并根据自己的需要对其进行化学处理。
这些需要是线粒体的需要。当突触前准备回收大部分未经修饰的递质时,就会出现第二次内部斗争,即线粒体的需求与囊泡释放机制对新鲜递质的需求之间的斗争。这些是游戏的简单基本规则,是突触生物计算机构建的基本逻辑。这是一个无限运行的捕获和释放程序,不断地被不同级别的反馈和前馈信号所修改,这些反馈和前馈信号由不同的发射器代谢物丰度所代表。为了充分理解为什么事情显然是这样设计的,我们必须评估这些产物对存在于我们每个不同传输系统两端的线粒体意味着什么。
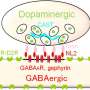
就gaba而言,我们已经观察到控制线粒体用于复制和DNA修复的核苷酸水平的一种重要活动。核苷酸在所有以g蛋白为基础的代谢受体上依次从其循环形式解锁,并进一步通过gaba转氨酶功能获得。gaba分流.
同样,血清素在新线粒体的形成中起着重要作用。反过来,含血清素的传播由线粒体内的核苷酸转运控制。
兴奋性递质谷氨酸直接参与运输还原物(NADH)到神经元线粒体通过与苹果-天门冬氨酸穿梭的相互连接。它通过谷氨酸-谷氨酰胺循环和线粒体谷氨酸脱氢酶直接穿梭于星形胶质细胞中。递质乙酰胆碱是由突触线粒体局部合成的胆碱和乙酰辅酶a合成的。它在突触间隙中被一种已知能产生显著细胞外质子电流的最快的酶降解。提供的胆碱也有助于脂肪酸代谢和膜脂合成,以建立和维护髓鞘。
儿茶酚胺能生态系统与关键线粒体电子传递功能的联系在这里完成了我们对所有主要神经递质家族的底层理解。虽然它不能直接解释在物种进化过程中多巴胺能回路中看到的高阶特征,但它很容易对突触结构的一些保守方面进行推测。例如,我们多巴胺能系统中的神经元的特征是无处不在的对称无脊椎动物“树突-树突”突触。相比之下,我们大脑的大多数其他区域部署了极化不对称连接,其中一边是囊泡,另一边是突触后密度。
区分我们对称突触区域的一件事是,我们已经将儿茶酚胺能传递器的朋友圈从无脊椎动物“章鱼胺”调整为多巴胺。随着新的连接体生成技术的出现像flybrain,我们现在可以读出与化学变化相对应的结构效应。事实上,现在做这件事很容易,机器可以通过编程自动完成。例如,人工智能识别算法可以在果蝇整个大脑的每个突触中找到每一个标志性的“多聚t条”释放位点。当我们可以为人脑做这些时,我们剩下的发射机神秘感将消失。
更多信息:Steven M. Graves等人。多巴胺代谢由单胺氧化酶线粒体穿梭激活电子传递链,自然神经科学(2019)。DOI: 10.1038 / s41593 - 019 - 0556 - 3
陈荣民等。多巴胺促进了自身的释放,自然神经科学(2019)。DOI: 10.1038 / s41593 - 019 - 0563 - 4
©2020科学欧宝app网彩X网络